
The Neural Substrate of Anxiety
 Photo courtesy of HealthlineOpens in new window
Photo courtesy of HealthlineOpens in new window
|
Anxiety, fear, aversion, and obsessive thinking are distinct symptomatic elements of anxiety disorders, but may reflect exaggerations of normal adaptive emotional responses. The most basic distinction in anxiety is between acute fear and anxious anticipation.
- Acute fear reflects the response to acute threat, activating the “fight or flight” response, and involves activation of a number of brain regions, including locus ceruleus and amygdale (Figure X-1).
- The latter is involved in the encoding of fearful memory and, hence, aversive conditioning. Therefore, as well as being involved in acute fear, the amygdale is also intimately involved in negative anticipatory expectation; that is, anxiety.
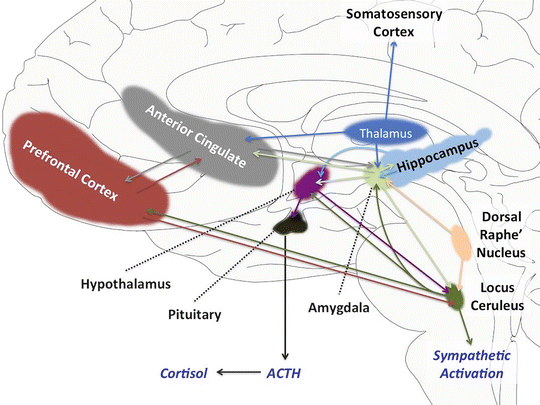 Source: The Medical Basis of Psychiatry (pp 145 - 168)Opens in new window
Source: The Medical Basis of Psychiatry (pp 145 - 168)Opens in new window
|
Acute fear activates the sympathetic nervous systemOpens in new window, producing peripheral manifestations such as tachycardiaOpens in new window, tachypneaOpens in new window, tremor, and diaphoresis. However, the perception of fear or anxiety involves the cortex, especially dorsolateral and orbital frontal regions.
The cortex not only consciously registers fear, but also responds with survival behaviors.
An important part of the connecting system between cortex and paralimbic structures is the cingulated gyrus, especially the anterior cingulated, which mediates the upward projection of fear- or anxiety-related information and downward regulatory signals, including both formal behavioral responses and more direct inhibitory activity.
Hence, the frontal cortex and cingulate are significantly involved in the regulation of both emotional and behavioral response to fear-inducing stimuli.
The anxiety-related brain regions normally exist in a relatively quiescent or inhibited state. Redundant inhibitory systems such as those related to gamma aminobutyric acid (GABA) and serotonin modulate the responsiveness of this system.
Additionally, neuropeptides such as corticotrophin releasing hormone, substance P, neuropeptide Y, cholecystokinin, and vasopressin, can have either activating or inhibiting effects. Therefore, this is a system that maintains a delicate balance of activation and inhibition.
This balance maintains optimal responsiveness (a desirable feature in a stress—threat response apparatus) while also inhibiting anxiety and fear in non-threatening situations. Alternatively, perturbation of one or more of these modulators may induce or maintain anxiety disorders.
Pharmacological treatments for anxiety disorders typically activate or augment that natural neuronal inhibitory mechanisms. GABAergic drugs such as the benzodiazepinesOpens in new window are a prime example.
GABA receptors exist in a complex with chloride channels. Chloride channels open in response to depolarization, propagating the action potential. By contrast, GABA receptors open chloride channels in a way that is not related to the propagation of action potentials, increasing the negative charge within neurons, and creating the inhibitory post-synaptic potential state. This inhibits subsequent action potentials until GABA is unbound and the chloride channels close, resulting in hyperpolarization. This decrease in neuronal firing appears to be related to the action of drugs that act on GABA receptors; GABA activation inhibits anxiety and can induce sleep.
There are GABA potentiating drugs such as baclofen and gabapentin that exhibit anti-anxiety properties. However, they are not commonly prescribed for anxiety. Receptors for the benzodiazepine drugs such as diazepam and alprazolam exist in the same GABA-chloride channel complex.
Benzodiazepines facilitate the action of GABA, which enhances and sustains the channels in the open position, producing a logner lasting effect than with GABA alone and, hence, reducing anxiety.
Serotonin from the raphé nuclei in the brainstem also serves an important regulatory role with regard to the structures that mediate anxiety.
SerotoninOpens in new window is a complex transmitter in the CNS, with multiple receptor subtypes that can have opposing functions. However, the inhibition of amygdala activity appears to occur primarily via serotonin 1A receptors.
Therefore, drugs that enhance activation of this receptor will produce inhibition of amygdala activity. These would include inhibitors of the serotonin transporter (typically referred to as selective serotonin reuptake inhibitors [SSRIs]) such as sertraline and paroxetine, and the serotonin 1A partial agonist buspirone.
As noted earlier, activation of the locus ceruleus, located in the brain stem, is a significant mechanism mediating anxiety, and is thought to be an important brain structure in the generation of acute anxiety.
Since anxiety disorders produce symptoms akin to acute anxiety, the locus ceruleus has been hypothesized to be important in the generation of panic attacks. SSRIs have an inhibiting effect on locus ceruleus firing, although only after chronic administration (typically 3 weeks).
The effect of serotonin at the locus ceruleus appears to be mediated via a different mechanism than in the amygdale. The serotonin 1A partial agonist buspirone actually enhances locus ceruleus firing activity. This may be related to the relative lack of effect of buspirone in panic disorder and reports of the drug actually inducing panic attacks.
Activation of the serotonin 2A type of receptors, on the other hand, inhibits locus ceruleus firing activity. Therefore, the net effect of blocking the reuptake of serotonin is to inhibit both amygdala and locus ceruleus activity. This may explain the broad effects of SSRIs on different anxiety disorders.
See also:
- Mineka S, Zinbarg R. A contemporary learning theory perspective on the etiology of anxiety disorders: it’s not what you thought it was. Am Psychol 2006; 61: 10 – 26.
- Stein MB, Goldin PR, Sareen, J, Zorrilla LT, Brown GG. Increased amygdale activation to angry and contemptuous faces in generalized social phobia. Arch Gen Psychiatry 2002;59: 1027 – 1034.
To be conscious of this state of affairs means, according to theory, that one entertains a second-order thought about the seeing—that is, the second-order thought represents the first-order seeing.
A weaker version does not require that one has to entertain the second-order thought, but only that one has to have the potential for having the second-order thought (Carruthers, 1996). That some such condition must be true can be seen from the following consideration:
“Could it ever be that I can genuinely claim that I am consciously aware of the chocolate being in the cupboard, but claim ignorance of the first-order mental state by which I behold this state of affairs—that is, by claiming that I have no clue as to whether I see, or just think of, or want the chocolate being in the cupboard?”
The important point of these conceptual analyses is that to be conscious of some fact requires some minimal concept of knowledge or of some perceptual state like seeing.
Unfortunately, there is no clear evidence when children understand a minimal state of this sort. There is some evidence of understanding (mother’s) emotional reactions and seeing (direction of gaze) in the first year of life (see Perner, 1991, chap. 6; Baldwin & Moses, 1996; Gopnik & Meltzoff, 1997, for summaries and discussion of problems of interpretation).
There is also some recent evidence that between 8 and 12 months children might be inferring people’s intentions to grasp an object from where that person looks (Spelke, Philips, & Woodward, 1995) and even between 5 to 9 months from how a person touches an object (seemingly intentional or accidentally).
And by 18 months (where children’s understanding of mental phenomena seems to flourish in general) children imitate people’s intended actions even when they observe a failed attempt (Meltzoff, 1955a) and they understand differences in preferences (e.g., that someone else can prefer cauliflower over biscuits, Repacholi & Gopnik, 1997).
Evidence that children distinguish their knowledge from ignorance is available at a relatively late age. Povinelli, Perilloux, and Bierschwale (1993) asked children to look for a sticker under one of three cups.
Children were first trained to look under the cup at which the experimenter had pointed. After some training even the youngest were able to do this.
When asked to look without the experimenter pointing, an interesting developmental difference emerged. Children older than 2 years and 4 months acted without hesitation when they knew which the cup the sticker was under, but hesitated noticeably when—in the absence of the experimenter’s poining—they had to guess where it was.
Interestingly this is also the age at which children start using the phrase “I don’t know” (Shatz, Wellman, & Silber, 1983). In contrast, children younger than that showed no comparable difference in reaction time. This may indicate that young 2-year-olds do not yet reflect on what they do and do not know.
So, theory of mind research is not yet able to give a guideline for when infants might develop explicit, conscious memories. Memory development may help out on this point.
Meltzoff (1985, 1995b) demonstrated that 14-month-old infants can reenact a past event (e.g., they imitate the experimenter leaning forward to touch a panel with forehead so that panel lights up) after several months. Recently this has been demonstrated in 11-month-olds with a delay of 3 months.
Since this is achieved from a brief observational period and does not require prolonged learning, and since patients with amnesia cannot do this (McDonough, Mandler, KcKee, & Squire, 1995), it is tempting to conclude that such enactment demonstrates explicit, conscious memory.
One should, though, keep in mind that delayed imitation that is based on a single event (third-person view) is not to be equated with a memory (knowledge) of that event as a single, past event (first-person view).
- Adapted from: The Oxford Handbook of Memory. Authored by ENDEL TULVING (ED.), Fergus I. M. Craik
