
Flowers
Flowers Anatomy and Functions
 A flowering apple tree | Credit: thisNZlifeOpens in new window
A flowering apple tree | Credit: thisNZlifeOpens in new window
Flowers are the organs of sexual reproduction in flowering plants. They are where fertilization, the fusion of the male sex cell in pollen and the famele sex cell in the ovule, takes place. |
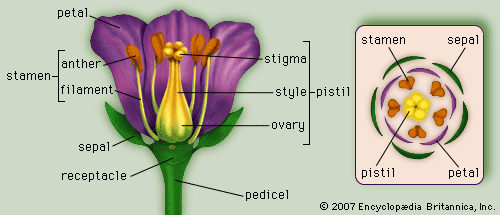 A typical structure of a monocotyledonous flower | Credit: BritannicaOpens in new window
A typical structure of a monocotyledonous flower | Credit: BritannicaOpens in new window
Although varying vastly from species to species, a “typical” flower normally is composed of sepals, petals, stamens, and pistils. |
These four floral organ types are arranged in whorls and are attached to a stem tip referred to as the receptable. The numbers of sepals, petals, stamens, and carpels (leaflike structures that comprise the body of the ovaries) are relatively constant for a species.
The number is typically five (or multiples of five) for dicots and three (or multiples of three) in monocots, although other numbers may occur (e.g., four sepals and four petals in the Cruciferae/Brassicaceae).
Sepals tend to appear leaflike and enclose the bud, and they may be fused at their bases. Collectively, the sepals are called the calyx. The petals, collectively the corolla, are usually showy, white, or colored, and are attractive to insects and other animals that could be potential pollinating agents.
Nectaries may also occur, usually at the base of the petals, and they normally secrete a sweet liquid known as nectar, which also tends to attract pollinators. Sometimes the petals are fused into a tubular, funnel-like or bellike shape, as in tobacco, petunia and Campanula, respectively.
When sepals and petals are showy and colorful but indistinguishable from one another, such as in Liliaceae members tulip and lily, they are called tepals, rather than sepals or petals. The calyx and corolla are collectively termed the perianth and when fused together (often with the stamen and ovary bases also), they are called the hypanthium.
The stamens are the male organs of the flower and consist of two parts, the pollen-bearing part called the anther and the usually slender stalk upon which the anther is born, called the filament.
The pollen is shed by the splitting, or dehiscence, of the anther (anthesis). Stamens of many plants may appear as highly modified structures that are petal-like in appearance called staminodes.
Flowers with numerous staminodes are the “double” flowers of many important horticultural species such as cultivated roses. Indeed, some botanists consider the normal petals of many flowers to have evolved from stamens and therefore to be staminodes.
The pistil is the female part of the flower and is comprised of one or more carpels, which are modified leaves bearing ovules along each edge. If the pistil has a single carpel, it is called a simple pistil and the carpel is folded lengthwise so that the ovules are protected and enclosed.
In ovaries with more than one carpel (a compound pistil), the carpels are fused along adjacent edges, thus forming a tubular structure, somewhat analogous to tubular forms of a corolla or calyx.
Whether simple or compound, a pistil is normally differentiated into three parts: the basal portion, or ovary, contains the ovules; the stigma is the upper portion and receives the pollen; and the style is the usually slender part of the pistil that connects the stigma to the ovary.
In compound pistils, the stigma may be split or lobed into as many parts as there are carpels — three for lily. The ovary chambers are referred to as locules. The placement of the ovules in the locules, or on the interior of the carpels, is termed placentation and is a useful tool in plant identification (see Figure YX-1).
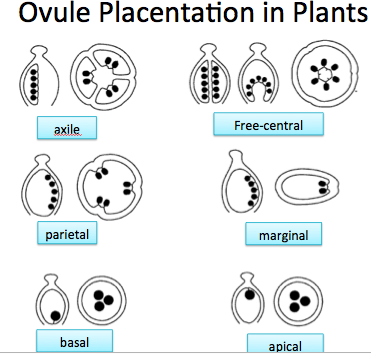 Figure YX-1. Ovules placentation | Credit: QuizletOpens in new window
Figure YX-1. Ovules placentation | Credit: QuizletOpens in new window
|
If all four kinds of floral organs (sepals, petals, stamens, and pistils) are present, the flower is said to be complete; if one or more floral organ types is missing, the flower is incomplete. The stamens and pistil are considered the essential parts and if both are present, the flower is considered to be a perfect flower; if either is absent, the flower is termed imperfect.
Perfect flowers are also called hermaphroditic (from the Greek mythological figure, Hermaphroditus, who was the son of Hermes and Aphrodite and was united when bathing into a single body with the nymph Salmacis). They are typical of flowers found in many angiosperm families.
If the stamens are absent, and only pistils present, the flower is said to be pistillate, and it is staminate if stamens are present and pistils absent.
A plant with both staminate and pistillate flowers is referred to as monoecious (literally “one house” meaning both sexes in one house); examples are oak, corn (maize), begonia, and squash.
Conversely, a dioecious species is one in which staminate flowers and pistillate flowers occur only on separate plants; examples are asparagus, holly (Ilex), Ginkgo, and papaya (Carcia papaya). If a plant has both perfect and staminate flowers, as in some Cucurbitaceae, it is said to be andromonoecious and if only pistillate flowers exist, the plant is gynoecious.
This latter character (gynoecy or all-femaleness) has been actively sought in cucumber breeding programs, resulting in numerous commercial gynoecious pickling cucumber cultivars that set their fruit in a concentrated fashion, thus facilitating once-over machine harvest of desirably sized fruits.
It is necessary, however, to include seeds of some stamen-bearing (andromonoecious) cultivars when planting a field of gynoecious cucumbers to provide a pollen source for adequate fruit set.
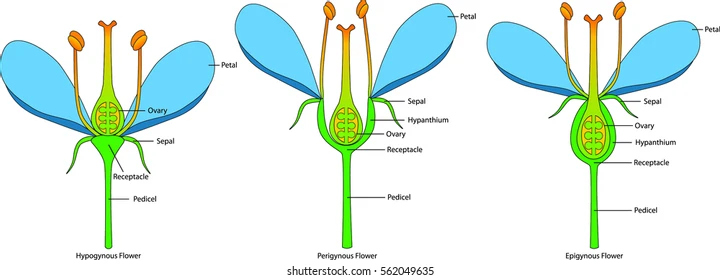 Figure YX-2. Ovary positioning | Credit: shutterstuckOpens in new window
Figure YX-2. Ovary positioning | Credit: shutterstuckOpens in new window
|
Position of the ovary is another useful parameter in classification and identification of plants. The types of ovary position are illustrated in Figure XY-2, and include hypogynous (hypo = below) wherein the origin of the bases of the sepals, petals and stamen is below, and distinct from, the ovary base (superior ovary).
A modification of this condition is also illustrated; the perigynous (peri = around or encircling) flower is still an example of a superior ovary, but the fused bases of sepals, petals, and stamens (hypanthium) surround the ovary, without fusing with the ovary as they do in the epigynous flower (epi = above).
In an epigynous flower, the hypanthium is fused to the ovary walls so that the sepals, petals, and stamens appear to arise from the top of the ovary, hence the term inferior ovary. Lily and raspberry flowers are hypogynous; cherry flowers are perigynous; and flowers of the members of the Cucurbitaceae are epigynous.
 Figure YX-3. Texas milkweed (An example of inflorescence) | Credit: BritannicaOpens in new window
Figure YX-3. Texas milkweed (An example of inflorescence) | Credit: BritannicaOpens in new window
|
The placement or arrangement of flowers on a stem can also vary greatly from species to species. Flowers may be borne singly (solitary flowers) or may be grouped in various clusters called inflorescences (see Figure YX-3). Solitary flowers and inflorescences are borne on stems called peduncles, the tip of which is the receptable.
The smaller stemlike structures bearing the individual flowers in an inflorescence are called pedicels. Each flower in an inflorescence is usually subtended by a bract.
Some specialized inflorescences are typical of certain taxa or groups, such as the umbel for the Umbelliferae (Apiaceae) and the spikelet found on the spikes, racemes, and panicles of the Graminae (Poaceae).
The highly membranous tough modified leaves known as glumes, lemma, and palea serve as the outer floral organs in such grass flowers. Like many other species of wind pollinated plants, these flowers tend to be inconspicuous and less showy.
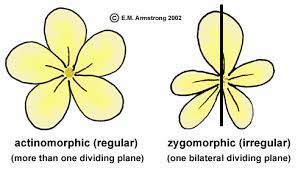 Figure YX-4. Flower symmetry | Credit: Waynes WordOpens in new window
Figure YX-4. Flower symmetry | Credit: Waynes WordOpens in new window
|
There are two general types of inflorescences based on their pattern of floral opening: the determinate, in which the apical bud opens first, followed sequentially by those below and working downward (e.g., cymes of strawberry) or indeterminate, typical of many other common inflorescence types including panicles and racemes.
Because botanists tend to think of fusion of floral organs and/or reduction in numbers as evolutionary advancements, species in the Compositae (Asteraceae) that have a head (capitulum) are considered more advanced than those species in which panicles are the norm. Regular flowers, radially symmetrical, are thought to be less advanced than irregular flowers, which are bilaterally symmetrical. (Figure YX-4)
You might also study other plant organs:
- Bailey, L.H., E. Z. Bailey, and the Staff of the L.H. Bailey Hortorium. 1976. Hortus Third: A Concise Dictionary of Plants Cultivated in the United States and Canada. MacMillan, New York.
- Bailey, L. H. 1949. Manual of Cultivated Plants Most Commonly Grown in the Continental United States and Canada, (rev. ed.). MacMillan, New York.
- Bidwell, R.G.S. 1979. Plant Physiology. (22nd ed.). MacMillan, New York.
- Devlin, R.M. and F. H. Witham. 1983. Plant Physiology. (4th ed.). PWS, Boston.
- Esau, K. 1977. Anatomy of Seed Plants. (2nd ed.). Wiley, New York.
- Heyward, H. E. 1938. The Structure of Economic Plants. MacMillan, New York.
- Lawrence, G. H. M. 1951. Taxonomy of Vascular Plants. MacMillan, New York.
- Leyser, O. and S. Day. 2003. Mechanisms in Plant Development. Blackwell, Oxford, UK.
- Mauseth, J. D. 1988. Plant Anatomy. Benjamin/Cummings, Menlo Park, CA.
- Norstog, K. and R.W. Long. 1976. Plant Biology. Saunders, Philadelphia.
- Ray, P.M., T. A. Steeves and S. A. Fultz. 1983. Botany. Saunders, Philadelphia.
- Salisbury, F.B. and C.W. Ross. 1978. Plant Physiology. Wadsworth, Belmont, California.
- Walton, P.D. 1988. Principles and Practices of Plant Science. Prentice Hall, Englewood Cliffs, N.J.
- Wilson, C. L. and W.E. Loomis. 1967. Botany, (4th ed.). Holt, Rinehart and Winston, New York.
