
Prefrontal Cortex (PRC)
The Prefrontal Cortex Contributions to Cognitive Control of Memory
To be adaptive, the long-term memory system must support retrieval of previously stored knowledge that has high utility given our current task and goals. In this framing, the problem of memory retrieval concerns balancing the recovery of useful information on the one hand against the inherent costs associated with retrieval itself (Anderson & Milson, 1989).
Central to striking this balance is cognitive control function (sometimes called executive function), or the ability to leverage abstract goals and contextual representations in order to adaptively influence retrieval and memory-based performance.
The Prefrontal Cortex (PFC) is necessary for cognitive control function, including during the cognitive control of memory. Whereas damage to medial temporal lobe (MTL) Opens in new window structures produces amnesia that catastrophically impairs the encoding of new information and retrieval of recently encoded information (Scoville & Milner, 1957), damage to the prefrontal cortex (PFC) results in more subtle memory deficits (Moscovitch, 1992; Stuss & Alexander, 2005).
For example, prefrontal cortex (PFC) patients are impaired in contexts that:
- require retrieval of specific information (i.e., source memory tasks; Janowsky et al., 1989; Swick, Senkfor, & Van Petten, 2006),
- reliance on retrieval strategies (Moscovitch & Melo, 1997),
- overcoming interference (Moscovitch, 1982; Squire, 1982; Winocur, Kinsbourne, & Moscovitch, 1981),
- ordering information at retrieval (Shimamura, Janowsky, & Squire, 1990), or
- retrieval with limited cue support (e.g., free recall; Janowsky, Shimamura, & Squire, 1989; Jetter et al., 1986; Stuss et al., 1994).
Neuroimaging studies have similarly implicated the prefrontal cortex (PFC) in relation to specific manipulations of cognitive control at retrieval (Badre & Wagner, 2007; Fletcher & Henson, 2001; Rugg & Wilding, 2000). Thus, prefrontal cortex (PFC) is a crucial component of the system that supports cognitive control of memory.
In general, the mechanism of cognitive control can be described in terms of a process of guided activation (Miller & Cohen, 2001), wherein contextual or goal information is maintained in working memory and thereby has the opportunity to provide a top-down influence on processing elsewhere.
Nevertheless, a central debate in the study of cognitive control concerns whether cognitive control is a unitary process or involves a diverse set of functionally distinguishable control processes (e.g., Cooper, 2010).
Mirroring the debate about the componentiality of cognitive control, the prefrontal cortex (PFC) is likely not a functionally homogenous structure supporting a unitary executive, but may contain distinct subsystems that support different forms of cognitive control. Neuroimaging studies have provided the primary evidence in favor of functional dissociations in prefrontal cortex (PFC) (Badre & Wagner, 2007; Simons & Spiers, 2003; Spaniol et al., 2009).
Though debate still exists as to whether there are undifferentiated “multiple-demand zones” within the PFC—such as within the mid-dorsolateral PFC (Duncan, 2010)—it seems now widely accepted that functional distinctions likely exist, such as between
- ventral and dorsal lateral PFC (Petrides, 2002; Simons & Spiers, 2003),
- rostral versus caudal PFC (Buckner, 2003; Race, Shanker, & Wagner, 2008),
- left versus right lateral PFC (Nolde, Johnson, & Raye, 1998; Tulving et al., 1994), and
- lateral PFC versus medial and subcortical systems (Kuhl et al., 2008; Scimeca & Badre, 2012).
Considerable controversy remains, however, regarding the validity of these distinctions and how to map them onto both individual experimental tasks and real-world behavior.
The cognitive control of memory is also likely componential, emerging from a set of interacting component processes. For example, cognitive control has the opportunity to influence retrieval performance in several ways, ranging from processes that structure inputs to the memory system (e.g., cue elaboration) to output control that monitors the outcome of retrieval and selects which representations are permitted to influence decision and action (Benjamin, 2007).
These and other distinct memory control processes could likewise be supported by different brain systems. Though research has only begun to refine understanding of these mechanisms and their neural correlates, several distinctions have been proposed regarding differential control processing in the prefrontal cortex (PFC) (e.g., Badre & Wagner, 2007; Nyberg, Cabeza, & Tulving, 1996; Simons & Spiers, 2003; Spaniol et al., 2009; Tulving et al., 1994).
Here we will focus on an example of functional specification within PFC related to cognitive control of memory retrieval. We will discuss the hypothetical specialization within ventrolateral PFC (vlPFC) between controlled retrieval and post-retrieval selection/monitoring operations. We will conclude by broadening the discussion of this distinction to consider the participation of these PFC subregions within distinct larger-scale functional networks.
Venterolateral PFC and the Two-Process Model
The left vlPFC refers to the broad region of lateral frontal cortex that is ventral to the inferior frontal sulcus and rostral to promotor cortex (Figure X). Investigation of the function of this region has long provided the strongest evidence in favor of functional specialization within PFC, from the classic studies of language impairment by Paul Broca (e.g., Broca, 1861) to early functional magnetic resonance imaging (fMRI) studies that distinguished subregions within left vlPFC related to the domain of verbal processing (e.g, the semantic versus phonological distinction; Poldrack et al., 1999).
| Figure X | Anatomical divisions of vlPFC | Credit: ResearchGate.net Opens in new window |
|---|
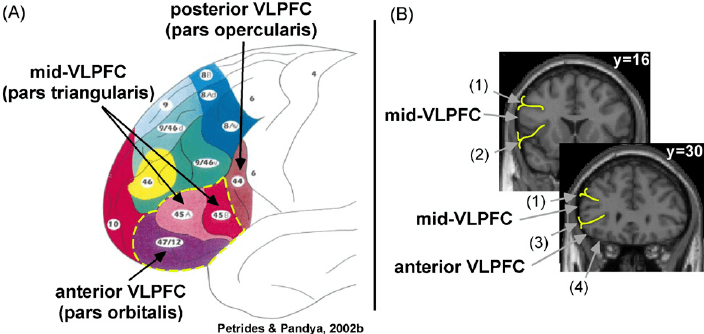 |
| Figure X | Anatomical divisions of vlPFC. (a) Schematic representation of the cytoarchitectonic divisions of the lateral PFC (adapted from Petrides and Pandya, 2002). Labels highlight the anterior vlPFC (parts orbitalis ( ~ Brodmann area [BA] 47)) and mid-vlPFC (pars triangularis (~ BA 45)). (b) Coronal slices from the Montreal Neurological Institute (MNI) canonical brain depict the anatomical boundaries that define mid-vlPFC and anterior vlPFC (reprinted with permission from Badre and Wagner, 2007). Labeled anatomical boundaries are (1) inferior frontal sulcus, (2) insular sulcus, (3) horizontal ramus of the lateral fissure, and (4) orbital gyrus. |
More recent work has focused on functional distinctions within this region as they relate to the cognitive control of memory. Specifically, recent work in our lab and in others has focused on a distinction between controlled retrieval, supported by anterior vlPFC, and post-retrieval selection, supported by mid-vlPFC (Badre and Wagner, 2007).
Here, we will consider the evidence for and against this potential distinction within the left vlPFC. To illustrate the distinction at the process level between controlled retrieval and selection, it is helpful to consider the analogy of searching for specific information on the Internet. For example, consider that you wish to find information about our lab.
First, you need to “hit” our link from the broad, latent associative structure of the web. To do this, you devise a particular keyword to put in your web browser. Of course, some keywords will be more effective than others. For example, searching for “Badre lab” is likely to produce our lab’s website as the top link. However, a less effective search, such as “science lab,” would make it unlikely that you would find the link to our lab without a prohibitive cost in browsing time.
Similar to this example, controlled retrieval refers to strategically guiding the activation of task-relevant information from its latent state. In human memory, controlled retrieval can progress by focusing on or elaborating effective cues and thereby increasing the likelihood that task relevant information is activated from memory.
From this perspective, one means of manipulating controlled retrieval experimentally is to reduce the strength of association between salient cues and target knowledge that would support automatic, cue-driven retrieval. In these cases, a top-down influence can aid in activating relevant knowledge from memory.
Importantly, however, it is very difficult to devise even a pair of keywords in a search engine that produce only a single web link. (In fact, it is rare enough that there is a hobby called “Googlewhacking”, whereby people try to achieve fame by finding pairs of keywords that produce only one hit through GoogleTM.) Thus, once we retrieve information into our browser, we “browse” or further select the links we want from this limited retrieved set.
Though human memory is different in important ways from the Internet, it is a similarly vast, associative structure that uses a form of priority, such as previous co-occurrence, to rank the likelihood that a given representation will be retrieved given a particular cue (Anderson and Milson, 1989; Griffiths, Steyvers, and Firl, 2007).
But, as on the Internet, this associative structure ensures that multiple representations will be retrieved given any cue, and that the highest-ranked representation may not be the one that is needed, given current goals and decision criteria. Thus, it is adaptive if a controlled retrieval system is complemented by an output control system that maintains current decision criteria and selects relevant items from among competitors in working memory Opens in new window.
The process of selecting from among retrieved information is termed post-retrieval selection. From this perspective, manipulations of response or decision criteria or varying the degree of competition among retrieved representations should affect post-retrieval selection.
Multiple lines of evidence support the involvement of the vlPFC in the cognitive control of memory. Functional neuroimaging studies, including using fMRI and positron emission tomography (PET), have repeatedly demonstrated greater activation in vlPFC under conditions of effortful or goal-directed retrieval, such as:
- controlling phonological and semantic representations (e.g., Gold et al., 2005),
- retrieving items with weak versus strong cue support (e.g., Badre et al., 2005),
- overcoming proactive interference (e.g., Ӧztekin and Badre, 2011) and,
- active inhibition of memories (e.g, Anderson et al., 2004).
Moreover, disruption of vlPFC due to neurological damage or disease decreased patients’ ability to select among competing information (Metzler, 2001; Thompson-Scrill et al., 1998). Similarly, intraoperative stimulation (Klein et al., 1997) or application of transcranial magnetic stimulation (Devlin, Mathews, and Rushworth, 2003; Gough, Nobre, and Devlin, 2005) disrupts performance when participants are required to retrieve semantic information.
Thus, in broad terms, vlPFC makes a necessary contribution to cognitive control of memory retrieval. However, an ongoing debate concerns the precise nature of vlPFC contributions to memory retrieval, and its functional organization in support of cognitive control.
Drawing on a wide range of declarative memory paradigms, including tests of both semantic Opens in new window and episodic memory Opens in new window, as well as retrieval during action selection, such as task switching, Badre and Wagner (2007) proposed that distinct subdivisions of the rostral left vlPFC support distinct controlled retrieval and post-retrieval selection processes, associated with the inferior frontal gyrus (IFG) parts orbitalis (~Brodmann area [BA] 47) and parts triangularis (~ BA 45), respectively. These subregions were termed anterior vlPFC and mid-vlPFC, respectively (refer back to Figure X ). We now briefly summarize the evidence that supports this distinction.
The two-process model proposes that anterior vlPFC is activated when memory must be searched in a goal-directed manner (i.e., controlled retrieval; Badre and Wagner, 2007). Accordingly, when bottom-up cues are insufficient to elicit activation of target knowledge (i.e., automatic retrieval), demands on controlled retrieval process increase.
Control can aid retrieval in these contexts by elaborating Opens in new window cues or generating retrieval plans that structure the input to the retrieval system and so make it more likely that relevant information will be retrieved.
| Figure Xz | Anatomical representations of mid-vlPFC and anterior vlPFC | Credit: ScienceDirect.com Opens in new window |
|---|
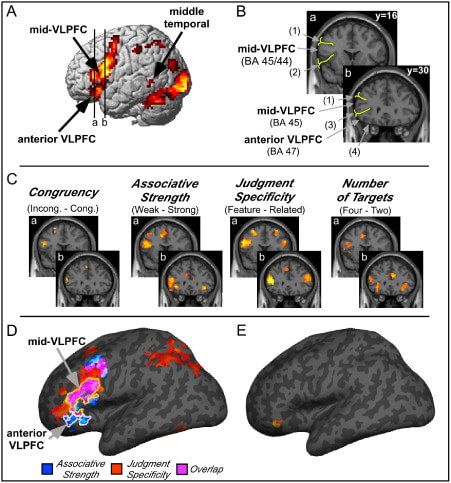 |
| Figure Xz | Results from manipulations of control during semantic retrieval provide evidence for the two-process model (reprinted with permission from Badre et al., 2005). (a) Contrasts of weak relative to strong associative strength (associative strength) (p < 0.001) and decisions of item similarity based on features (e.g., color) relative to general semantic relatedness (feature specificity) (p < 0.001). (b) Contrasts of associative strength (blue) and feature specificity (red) and their overlap (purple) are rendered on an inflated MNI canonical surface. Anterior vlPFC was sensitive to associative strength, whereas mid-vlPFC was sensitive to both associative strength and feature specificity. |
Consistent with his hypothesis, anterior vlPFC is consistently activated during semantic retrieval tasks in which the association between available cues and target knowledge is weak. For example, deciding that “candle” is semantically related to “flame” is easier and requires less controlled retrieval than deciding that “candle” is related to “halo,” because the association between candle and halo is weak relative to the association between candle and flame. Thus, experiments that manipulate associative strength, based either on pre-experimental norms (Badre et al., 2005; Wagner et al., 2001) or on associations learned during the experimental session (Danker, Gunn, and Anderson, 2008), consistently show greater activation in anterior vlPFC under weak relative to strong associative strength conditions (Figure Xz Opens in new window).
In a way similar to high cue-target association strength, anterior vlPFC shows repetition suppression effects accompanying the increased semantic fluency that follows repetition of an item during a semantic memory task, even when the decision/response level effects are not repeated (Race, Shanker, and Wagner, 2008).
According to the two-process model, mid-vlPFC is activated under conditions in which multiple items are retrieved from memory, but only a subset must be selected for further processing (i.e., post-retrieval selection; Badre and Wagner, 2007). As described above, automatic and controlled retrieval processes can result in the recovery of multiple representations. Thus, post-retrieval selection is needed to resolve competition among the multiple retrieved representations, and to permit selected representation to guide decision and action.
Support for mid-vlPFC and post-retrieval selection comes from several sources. First, mid-vlPFC shows greater activation when participants are asked to decide if two items (e.g., “apple” and “blood”) are similar along a particular dimension, such as color, relative to deciding whether they are generally semantically related to one another regardless of dimension (feature specificity effect) (Badre et al., 2005; Thompson-Schill et al., 1997) (refer back to Figure Xz Opens in new window).
This difference is thought to arise because making the decision along a particular task-relevant dimension requires focusing attention only on the retrieved details relevant to the decision and ignoring any other properties. Notably, anterior vlPFC does not show a difference between specific and general decision conditions (Badre et al., 2005)
Proactive interference (PI) occurs when a prior learned association automatically elicits retrieval of information that competes with a current retrieval task (Anderson and Neely, 1996; Postman and Underwood, 1973). PI during short-term item recognition has consistently been associated with increases activation in mid-vlPFC (Badre and Wagner, 2005; Postle and Brush, 2004; Postle, Brush, and Nick, 2004). PI during short-term item recognition does not consistently produce activation increases in anterior vlPFC. However, as discussed above, other manipulations of PI have been associated with anterior vlPFC activation (Ӧztekin and Badre, 2011).
During lexical decision, an unexpected target produces an interference effect above a neural baseline. This interference effect is thought to be due to competition between information retrieved during preparation for the target and the information that must be retrieved upon encountering the unexpected target. Competition of this type during lexical decision is associated with increased activation in mid-vlPFC.
By contrast, anterior vlPFC shows priming effects consistent with the reduced retrieval demands (Gold et al., 2006). Thus, across these examples, it appears that mid-vlPFC is critical under conditions of competition, presumably when there is a demand to select relevant information for further processing. By contrast, anterior vlPFC is not consistently activated under these circumstances.
Importantly, attempts to directly dissociate anterior and mid-vlPFC are complicated by the fact that, akin to our Internet search analogy, any process of retrieval, be it controlled or automatic, holds the potential for competition. Thus, similar to anterior vlPFC, mid-vlPFC often shows increased activation under conditions requiring controlled retrieval (Badre et al. 2005; Wagner et al., 2001).
And so, though single dissociations are sometimes observed (e.g., Danker, Gunn, and Anderson, 2008), double dissociations are less common. However, if one pits competition against associative strength, it is possible to dissociate these regions. For example, when the number of retrieval cues is small (low overall retrieval) but associative strength is weak, there will be more demands on controlled retrieval and selection.
By contrast, when the number of retrieval cues is large (high overall retrieval) but associative strength is high, this puts greater demands on selection than controlled retrieval. Consistent with this prediction, Badre et al., (2005) directly pitted number of available retrieval cues against associative strength and produced activation in anterior vlPFC but not mid-vlPFC. Thus, crossing the number of retrieval cues (i.e., increasing retrieval demands) with associative strength dissociates anterior from mid-vlPFC; and when this is taken together with the feature specificity described above, this produces a region-by-effect interaction, dissociating anterior and mid-vlPFC (Badre and Wagner, 2007).
In summary, there is evidence both across and within studies for dissociable functions between anterior vlPFC and mid-vlPFC during cognitive control of memory, and these functions can be characterized as controlled retrieval and post-retrieval selection respectively.
Nevertheless, there have been challenges to the two-process model. These have included formal theoretical arguments about whether two processes are required to achieve controlled retrieval and selection function, as opposed to a single-process model that can support both functions (Danker, Gunn, and Anderson, 2008; Thompson-Schill and Botvinick, 2006). These models make clear that a single process could achieve these two functions. However, it would seem difficult for a single-process model to account for the empirical dissociation between these processes.
There has also been some debate about the nature of the relationship between anterior vlPFC and controlled retrieval, and whether the manipulation of associative strength actually reflects the domain of information being retrieved, such as retrieval of abstract semantics (Goldberg et al., 2007). Again, however, a strictly domain-based account appears too difficult to reconcile with the broader data supporting the controlled retrieval hypothesis, such as the observation of activation in anterior vlPFC when retrieving weak, arbitrary paired associations (Danker, Gunn, and Anderson, 2008).
See also:
- The Human HippocampusOpens in new window
- Role of The Hippocampus In RetrievalOpens in new window
- Role of the Parietal Cortex in RetrievalOpens in new window
- Role of the Prefrontal Cortex in RetrievalOpens in new window
- Role of the Amygdala in EmotionOpens in new window
- Anterior Temporal Regions: Amodal Storage of Semantic InformationOpens in new window
- Medial Temporal Lobe (MTL) Contributions to MemoryOpens in new window
Cited references in the book include:
- Allan, K., Wilding, E.L., and Rugg, M.D. (1998). Electrophysiological evidence for dissociable processes contributing to recollection. Acta Psychologica, 98 (2-3), 231-52. doi: 10.1016/S0001-6918(97)00044-9.
- Anderson, J. R., and Milson, R. (1989). Human memory: an adaptive perspective. Psychological Review, 96 (4), 703-719. doi: 10.1037/0033-2595X.96.4.703.
- Anderson, M.C., and Neely, J.H. (1996). Interference and inhibition in memory retrieval. In Memory Handbook of Perception and Cognition (ed. E. L. Bjork and R. A. Bjork). San Diego, CA: Academic Press, pp. 237-313.
- Anderson, M.C., Ochsner, K.N., Kuhl., et al., (2004). Neural systems underlying the suppression of unwanted memories. Science, 303 (5655), 232-235. doi: 10.1126/science.1089504.
- Badre, D., Poldrack, R.A. Pare-Blagoev, E.J., et al., (2005). Dissociable controlled retrieval and generalized selection mechanisms in ventrolateral prefrontal cortex. Neuron, 47 (6), 907-918. doi: 10.1016/j.neuron.2005.07.023.
- Badre, D., and Wagner, A.D. (2005). Frontal lobe mechanisms that resolve proactive interference. Cerebral Cortex, 15 (12), 2003-2012. doi:10.1093/cercor/bhi075.
- Badre, D., and Wagner, A.D. (2007). Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia 45 (13), 28883-2901. doi: 10.1016./j.neuropsychologia.2007.06.015.
- Barredo, J., Ӧztekin, I., and Badre, D. (2013). Ventral fronto-temporal pathway supporting cognitive control of eipisodic memory retrieval. Cerebral Cortex, published online October 31, 2013. doi: 10.1093/cercor/bht291.
- Benjamin, A.S. (2007). Memory is more than just remembering: strategic control of encoding, accessing memory, and making decisions. In The Psychology of Learning and Motivation, Volume 48: Skill and Strategy in Memory Use (ed. A. S. Benjamin and B. H. Ross). New York, NY: Elsevier, pp. 175-223.
- Blumenfeld, R.S., and Ranganath, C. (2007). Prefrontal cortex and long-term memory encoding: an integrative review of findings from neuropsychology and neuroimaging. Neuroscientist, 13 (3), 280-291. doi: 10.1177/1073858407299290.
- Ranganath, C. (2010). A unified framework for the functional organization of the medial temporal lobes and the phenomenology of episodic memory. Hippocapus, 20 (11), 1263-1290. doi: 10.1002/hipo.20852.
- Ranganath, C. and Rainer, C. (2003). Neural mechanisms for detecting and remembering novel events. Nature Review Neuroscience, 4 (3), 193-202. doi: 10.1038/nrn1052.
- Schacter, D.L, and Slotnick, S.D. (2004). The cognitive neuroscience of memory distortion. Neuron, 44 (1), 149-160. doi: 10.1016/j.neuron.2004.08.017.
- Scoville, W.B., and Milner, B. (1957). Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery, and Psychiatry, 20 (1), 11-21. doi: 10.1136/jnnp.20.1.11.
- Tulving, E. (1985). Memory and consciousness. Canadian Psychology/Psychologie Canadienne, 26 (1), 1-12. doi: 10.1037/h0080017.
- Vargha-Khadem, F., Gadian, D.G., Watkins, K.E., et al. (1997). Differential effects of early hippocampal pathology on episodic and semantic memory. Science, 277 (5324), 376-380. doi:10.1126/science.277.5324.376.
- Yonelinas, A.P. (2002). The nature of recollection and familiarity: a review of 30 years of research. Journal of Memory and Language, 46 (3), 441-517. doi: 10.1006/jmla.2002.2864.
- Yonelinas, A.P., and Jacoby, L.L. (2012). The process-dissociation approach two decades later: convergence, boundary conditions, and new directions. Memory and Cognition, 40 (5), 663-680. doi:10.3758/s13421-012-0205-5.
- Yonelinas, A.P., and Parks, C.M. (2007). Receiver operating characteristics (ROCs) in recognition memory: areview . Psychological Bulletin, 133 (5),800-832. doi: 10.1037/0033-2909.133.5.800.
